Как рыбы видят в темноте. Зрение пресноводных рыб. Особенности органов чувств у костистых и хрящевых
Стопроцентной уверенности в том, как именно протекает жизнь под поверхностью воды, у нас нет. О том, как реагирует та или иная рыба на различные раздражители, каким образом она отыскивает приманку и что останавливает ее от решительной поклевки, мы судим косвенно — по результатам рыбалки, наличию-отсутствию «хваток» и сходов и т. д., и т. п.
Для того, чтобы эффективно применять свой рыболовный опыт в противостоянии с обитателями наших водоемов, современный рыболов- любитель или спортсмен обязан обладать немалым багажом знаний, полученных благодаря неоднократным личным наблюдениям или почерпнутых из достоверных научных источников.
В настоящей статье мы продолжаем разговор об органах чувств рыб и их неравнозначной роли в жизни подводных обитателей (см. «СР» №№ 2 и 8 за 2002 г., № 2 за 2003 г. и № 2 за 2004 г.).
Об органах чувств рыб
В истории развития человеческой цивилизации особое внимание изучению рыб начали уделять в IV веке до н. э. Фактически ихтиология как наука о рыбах началась с Аристотеля (384-322 гг. до н. э.), который сделал первые попытки классифицировать огромное разнообразие обитателей царства Нептуна и описывал биологию и анатомию многих видов рыб.
За две с половиной тысячи лет рыб изучили достаточно подробно, но естествоиспытатели II-XIX-го веков, описывающие в своих научных трудах подводных жителей рек, морей и океанов, были искренне уверены в том, что рыбы — это очень примитивные, глупые существа, которые не обладают ни слухом, ни осязанием, ни даже какой-либо памятью. Кстати, эти, в корне неверные, воззрения сохранялись в научной среде вплоть до 1940-х годов.
В настоящее время практически любой «литературно подкованный» рыболов, не говоря уже об ученых-ихтиологах, знает, для чего у рыб существует боковая линия, могут ли рыбы слышать или обонять, с помощью чего они отыскивают корм или чувствуют приближение хищника…
Общеизвестно, что органы чувств или, как принято их сейчас называть — сенсорные системы, дают возможность живому организму воспринимать разнообразную информацию об окружающем мире, а также сигнализировать о внутреннем состоянии самого организма.
Органы чувств рыб способны:
Воспринимать электромагнитные поля в видимой (зрение) и инфракрасной (температурная чувствительность) областях спектра;
Ощущать механические возмущения, или звуковые волны (слух),
Чувствовать силу тяжести (вестибулярная и гравитационная чувствительность) и механическое давление (осязание);
Распознавать разнообразные химические сигналы — восприятие веществ в жидкой фазе (вкус) и в газовой фазе (обоняние).
К сенсорным системам рыб можно отнести зрительную, слуховую, вкусовую, обонятельную, осязательную, электрорецепторную сенсорные системы, а также сейсмосенсорную систему, представленную боковой линией, общее химическое чувство.
К одним из самых значимых органов чувств у животных относится зрение — это способность воспринимать электромагнитные поля в видимой области спектра.
При помощи зрительных анализаторов рыбы ориентируются в пространстве, находят пищу или избегают хищников, занимают соответствующие экологические ниши, визуально оценивая характер зрительного окружения (Beur, Heuts, 1973).
Популярно о строении глаза рыб
Рыбы видят (воспринимают свет) в водной среде при помощи глаз и особых светочувствительных почек. Особенности видения рыб под водой обусловлены прозрачностью вод, их вязкостью и плотностью, глубиной, скоростями течений, способом жизни и питания.
По сравнению с наземными животными и человеком, рыбы более близоруки. Роговица их глаз плоская, а хрусталик шаровидный. Именно его форма и обуславливает близорукость у рыб. У многих рыб хрусталик может выступать из отверстия зрачка, благодаря чему увеличивается поле зрения.
Вещество хрусталика такой же плотности как и вода, в результате свет, проходя через него, не преломляется и на сетчатке глаза получается четкое изображение.
Сетчатка глаза (внутренняя оболочка) имеет сложное строение, состоит из четырех слоев: пигментного, светочувствительного (так называемые палочки и колбочки) и двух слоев нервных клеток, дающих начало зрительному нерву.
Роль палочек — функционирование в сумерках и ночью, причем они нечувствительны к цвету. При помощи колбочек рыбы воспринимают различные цвета.
Зрачок практически у всех видов неподвижен, однако камбалы, речной угорь, акулы и скаты в состоянии его сужать и расширять, увеличивая остроту зрения.
Особенности зрения у разных рыб
У большинства рыб движения глаз скоординированы, только у некоторых (зеленушка, калкан, морской язык и др.) они могут двигаться независимо друг от друга. У хищных рыб глаза наиболее подвижны.
У наших морских и пресноводных рыб органы зрения — глаза — расположены по бокам головы, причем каждый глаз видит свое поле зрения. Такое зрение называется монокулярным. Спереди монокулярное зрение каждого глаза перекрывается, появляется зона бинокулярного зрения. Угол бинокулярного зрения у рыб очень мал — не более 30?.
Известный американский ученый Роберт Вуд показал, как рыбы могут видеть из воды. По законам преломления световых лучей, предметы, находящиеся на суше, кажутся рыбе выше, чем на самом деле. Если смотреть из воды в сторону берега под углом к вертикали больше чем 45°, то из-за полного внутреннего отражения от поверхности воды наблюдателю (рыбе) становятся видны объекты (рыболов). Стоящий на берегу рыболов представляется ей висящим в воздухе и четко различимым, но сидящего человека рыба не заметит, так как под малым углом наклона лучей к горизонту (менее 45?) наземные объекты ей невидимы.
Подавляющее большинство пресноводных рыб видят максимум на 1 м. В прозрачной воде (например, в наших водохранилищах зимой) рыбы практически могут видеть на расстоянии 10-12 м, однако четко различают предметы, их форму, цвет в пределах 1-1,5 м. При аккомодации глаза с передвижением хрусталика глаз настраивается на расстояние, не превышающее 15 метров. Это предел дальности зрения рыб.
Согласно экспериментальным исследованиям, речной окунь в состоянии видеть предмет величиной 1 см на расстоянии около 5,5 метров. При уменьшении размеров предмета в 10 раз расстояние видения его хищником пропорционально уменьшалось — окунь видел предмет за 55 см. Крохотный объект величиной 0,1 мм хищник видел только за 5,5 см.
Ихтиологи различают светолюбивых (дневных) и сумеречных рыб. У дневных видов в сетчатке глаза палочек немного, зато колбочки большие. Эти рыбы (щука, плотва, голавль, жерех и др.) хорошо различают цвета — красный, синий, желтый, белый. У сумеречных рыб (судак, налим, сом,) в сетчатке находятся только палочки, и, следовательно, различать цвета и их оттенки они не в состоянии.
Глаза как орган зрения хорошо развиты у светолюбивых рыб (щука, чехонь, красноперка) и некоторых сумеречных видов (лещ, ерш, густера, налим). У других сумеречных рыб (придонных) — карпа, карася и линя — глаза развиты хуже (Протасов, 1968). В связи с этим у светолюбивых рыб ориентация и поиск в пространстве, питание могут осуществляться преимущественно с помощью зрения, а у сумеречных — главным образом благодаря органам осязания и других сенсорных систем.
У пелагических планктофагов (белый толстолобик, чехонь) поиск пищи осуществляется практически полностью благодаря зрению.
Способность рыб различать цвета. Дневные рыбы достаточно хорошо различают цвета, по крайней мере, спиннингисты об этом знают, применяя при разной освещенности белый виброхвост или бело-красный твистер в охоте на щуку или окуня. Черноморская хамса на фоне сине-зеленой воды различает (видит) сети разной окраски на следующем расстоянии: сине-зеленые — 0,5-0,7 метров; темно-синие — 0,8-1,2 м; темно-коричневые — 1,3-1,5 м; серые или черные — 1,5-2,0 м; белые (неокрашенные) — 2,0-2,5 м.
Сумеречные и ночные рыбы, как было отмечено выше, различать цвета не в состоянии, поэтому рыболовы-спортсмены и любители при экспериментировании с приманками должны уделять особое внимание не цвету приманки, а ее поведению (лобовому сопротивлению, шумовым характеристикам).
Применение специально ярко окрашенных приманок для ловли сумеречных хищников (тех же судака или сома) автору представляется неоправданным, так как эта рыба реагирует не на цвет некоего «Предатора», а только на его гидродинамические качества, корректируя предстоящий бросок видением (благодаря отличному сумеречному — черно-белому — зрению) абриса приманки. Причем чем ярче ее силуэт на фоне усеянного камнями дна (белое — на черном, флуоресцентное на черном), тем большее количество хваток и поимок хищника отметит спиннингист при применении одинаковых приманок, но разных расцветок. И снова решающее для броска судака значение будет иметь белый или желтый цвет приманки, а уж никак не фиолетовые, например, разводы на зеленом фоне воблера (если, конечно, это не супернеотразимая, гремяще-звенящая модель)…
Зрительное восприятие рыбами движений. Российские ученые исследовали способности зрительного аппарата рыб восприятия движения. Для этого наблюдали за оптомоторной реакцией рыб на последовательно движущиеся полосы или детали обстановки в течение 1 секунды (определение величины оптических моментов). Были получены следующие результаты.
Оптический момент у верховки и карася составил 1/14 — 1/18 секунды, щуки и линя — 1/25 — 1/28 с, леща и окуня — 1/55 с. Рыбы, имеющие оптические моменты от 1/50 до 1/67 с, способны вдвое детальнее воспринимать одно и то же движение, чем человек, а рыбы, имеющие оптический момент 1/10 — 1/14, — вдвое менее детально.
Тонкое восприятие движения зрительным аппаратом рыб позволяет жертвам уловить начальный момент броска и ускользнуть от хищника. Для мирных рыб сигналом предстоящего броска хищника являются подергивание и вибрирование спинных и грудных плавников, а также всего тела охотника, улавливаемые глазом потенциальной жертвы (Протасов, 1968).
Сытые и утомленные рыбы имеют слабо выраженную оптомоторную реакцию (реакцию на движение), а голодные и хорошо отдохнувшие — сильно выраженную реакцию.
Органы чувств рыб в пищевом поведении рыб
Представляют интерес для рыболова также и экспериментально полученные и проверенные в естественных условиях результаты поочередного функционирования органов чувств рыб при поиске ими кормовых объектов.
Во время «свободного поиска», когда расстояние до кормового объекта превышает 100 м, у рыб «работает» только обоняние, остальные сенсорные системы не задействованы. При приближении к источнику «вкусного» запаха от 100 до 25 м к обонянию подключается слух. На расстоянии от 25 до 5 м рыба пытается найти корм при помощи обоняния, зрения и слуха.
Когда до пищи остается «рукой подать» (от 5 до 1 м), рыба в первую очередь пользуется зрением, затем обонянием и слухом. На расстоянии от 1 до 0,25 м в поиск вовлекаются одновременно зрение, слух, боковая линия, обоняние, наружная вкусовая чувствительность (ощупывание грунта усиками, касания губами, рылом, даже плавниками).
Когда еда «под носом» и расстояние до нее не превышает 0,25 м, рыба «включает» практически все органы чувств: зрение, боковую линию, электрорецепцию, наружную вкусовую чувствительность, общее химическое чувство, осязание. Их совместная работа быстро приводит к обнаружению рыбой корма.
Поведение хищных рыб в зависимости от особенностей зрения
По отношению к периоду наибольшей пищевой активности применяют такое разделение хищных рыб: окунь — сумеречно-дневной хищник, щука — сумеречный, судак — глубокосумеречный.
Окуни-ихтиофаги и щуки питаются круглосуточно: днем охотятся за добычей из засады, в сумерках и на рассвете выходят на открытую воду и преследуют жертв. «Сумеречное» питание хищников происходит при освещенности от сотен до десятых долей люксов (вечером) и наоборот (утром). В этот период у окуня и щуки функционирует дневное зрение с максимальной остротой и дальностью видения, а плотные стаи рыб-жертв начинают распадаться, обеспечивая удачную охоту хищникам. С наступлением темноты отдельные рыбешки рассредоточиваются по акватории, верховка и уклейка при падении освещенности ниже 0,01 лк опускаются на дно и замирают. Охота хищных рыб прекращается.
В предутренние часы при освещенности от десятых долей до сотен люксов «избиение младенцев» продолжается до момента, когда рыбы-жертвы образуют плотные оборонительные стаи.
Согласно исследованиям ихтиологов, летом продолжительность утреннего питания хищников достигала 3 часов, вечернего — 4 часа и ночного (судак) — 5-6 часов.
Судак может пользоваться зрением в тех условиях, когда другие рыбы видеть не могут. Сетчатка глаза хищника содержит сильно отражающий свет пигмент — гуанин, который увеличивает ее чувствительность. Охота судака за мелкими стайными рыбами наиболее успешна при глубоко сумеречной освещенности — 0,001 и 0,0001 лк.
Осенью, в пасмурную и дождливую погоду, когда освещенность изменяется незначительно, молодь мирных рыб образует разреженные оборонительные стаи и хищники могут успешно охотиться на протяжении всего дня, а не только в сумерках. Происходит так называемый «осенний жор» хищника.
Подмечена интересная особенность охоты щуки и окуня на свету и при высокой прозрачности воды. В дневное время эти рыбы выступают как типичные хищники-засадчики: при неудачном захвате добычи из засады они не преследуют ее, чтобы не отпугнуть других потенциальных жертв от места охоты. Те районы, где затаился хищник, обнаруживший азартом свое место укрытия, стайки рыб обходят стороной. Поэтому днем щука или окунь делают четко выверенный и точный бросок только при возможности 100%-го захвата добычи. Решающую роль в удачном броске играет зрение.
Таким образом, зная об особенностях и возможностях зрительного восприятия рыб, рыболовы получают возможность осуществлять на водоеме целенаправленный поиск будущего подводного «спарринг-партнера». Знание сильных и слабых сторон противника (читай — возможностей зрения рыб в морской и пресной воде, днем и в сумерках), надеюсь, помогут многочисленным поклонникам рыбной ловли выходить победителем из этой увлекательнейшей и честной схватки…
Долго считалось, что у рыб нет цветового зрения, но уже лет 60 — 70 как доказано, что за исключением немногих донных и глубоководных видов, все остальные рыбы цвета вполне различают, и в некоторых отношениях даже лучше нас. Например, они видят ультрафиолетовые лучи и различают поляризованный свет, а мы — нет. Но есть у рыб и слабые стороны. Если мы хорошо различаем цвета всего видимого солнечного спектра, то рыбы — только его средней части. Края спектра для них представляются в одном цвете. Что это значит? Это значит, что, например, треска, пикша и зубатка не способны увидеть разницу между цветами левой части спектра — фиолетовым, синим, голубым и зеленым. Для них все они — один и тот же цвет. То же самое и с цветами правой части, оранжевым и красным. Фактически цветовое зрение у этих видов есть только для узкого среднего интервала цветов — они различают зеленый, желтый и оранжевый. Все это не означает, конечно, что в той области спектра, где рыбы дальтоники, они вообще не видят разницы между цветами. Они различают их по яркости, Как мы, например, видим разницу между светло-серым и серым. Так что, размышляя о цветовой гамме приманок, нужно понимать, что рыбы будут видеть их иначе, чем мы сами.
Вода часть падающего на ее поверхность света отражает, а что проходит — фильтрует. Это означает, во-первых, что под водой всегда меньше света, чем на воздухе, и, во-вторых, что он другой по своему составу. В результате на определенной глубине света не остается вообще, наступает полный мрак. Но при этом лучи разной длины волны поглощаются по-разному, одни исчезают на меньшей глубине, другие — на большей. В чистой морской воде быстрее всего поглощаются длинноволновые цвета — красный и оранжевый. Они не видны уже на глубине 5-8 метров. Затем исчезают желтый и значительно позже зеленый и синий. Такое выборочное поглощение имеет очень интересные последствия. Как будет выглядеть на глубине 10 метров красная приманка? На суше она отражает волны красного цвета (поэтому мы ее и видим красной), а все остальные поглощает. На 10-метровой глубине, как мы только что выяснили, красных лучей уже нет. Значит, нашей приманке там просто нечего отражать. Как она будет выглядеть? Правильно, она будет черной. Самые неожиданные превращения могут происходить с приманками, в окраске которых имеется белый цвет. Белыми, как говорилось, выглядят предметы, которые отражают сразу все семь цветов спектра. Если же какие-то цвета отсекаются в результате поглощения водой, то они, естественно, не падают и на приманку и, следовательно, не отражаются от нее. В результате из белого «букета» исключается часть лучей, и белое становится цветным.
Но это все для случая чистой и прозрачной морской воды. В пресных водах дело обстоит сложнее. Наши приманки могут самым решительным образом менять свою «боевую раскраску» в зависимости оттого, на каком водоеме мы собираемся ловить на них рыбу. Но и это еще не все. Характер поглощения света может быть разным на разных участках одного и того же водоема и даже на одном и том же участке в разные часы суток.
Выводы: Для очень многих условий ловли цвета приманок вообще не играют никакой роли. Но на самом деле это не совсем так. Есть несколько моментов, которые смягчают этот приговор цвету. Глаза рыб наиболее чувствительны к свету определенной длины волны, то есть, определенного цвета. Для пресноводных рыб это красно-оранжевый цвет при дневной освещенности, и зеленый в сумерках (показано для линя, леща, карася, окуня, налима и басса). Другими словами, при сумеречном освещении зеленые предметы рыбы видят лучше и с большего расстояния, чем предметы других цветов. То же самое справедливо для красно-оранжевых предметов при свете дня.
Подводя итоги всему сказанному, можно сформулировать главный и, на первый взгляд, парадоксальный вывод. Цвет рыболовной приманки не есть что-то заданное и неизменное. На самом деле, то, как его воспринимает рыба, зависит от свойств воды водоема, в который эта приманка забрасывается, и от освещенности, в которую она при этом попадает.
Изменения окраски тела рыб связано с тем, что рыбы приспосабливаются к тем условиям, в которых они живут, окраска их тела становится похожа на цвет грунта, либо приобретают своего рода «камуфляжную» окраску, если обитают среди водных растений. Надводный мир по сравнению с животными, живущими на суше, рыбы видят несколько иным. Если смотреть вертикально вверх, то рыбы видят все без искажения, а если под углом в сторону, то из-за преломления луча зрения и двух сред – воздуха и воды, картинка искажается.
Зрение у рыб . У рыб максимальная видимость в прозрачной воде не превышает 10 – 12 метров, это все по тому, что оптические свойства воды не позволяют далеко видеть. Расстояние видимости может и сокращаться, причиной этому может послужить: цвет воды, мутность воды, освещенности и т.д. На расстоянии не более 2 метров рыбы видят предметы наиболее четко. Лучше всех видят хищники, предпочитая день, обитающие в прозрачной воде, — форель, хариус, щука, жерех. У некоторых рыб, питающихся планктоном и донными организмами (сом, лещ, угорь, налим, судак и др.) в сетчатке глаза имеются такие светочувствительные элементы, которые способные воспринимать слабые световые лучи. Благодаря этим элементам эти рыбы довольно хорошо видят и в темноте.
Угол зрения рыб устроен таким образом : они могут видеть предметы в зоне около 150° по вертикали и до 170° по горизонтали. Из воды в воздухе рыба видит предметы как бы через круглое «окно», ограниченное углом зрения порядка 97°. Соответственно, если рыба подплывет ближе к поверхности, то «окно» будет становиться все меньше и меньше.
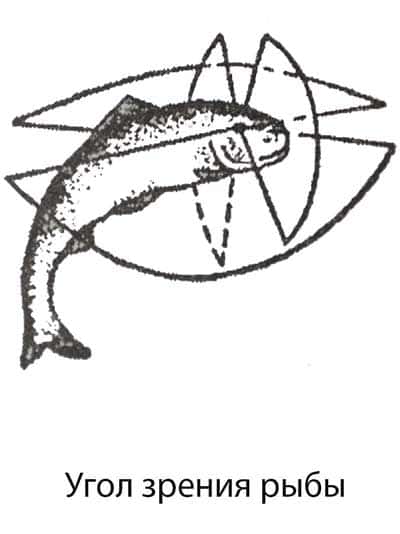
Видит ли рыба рыболова
Вблизи берега рыба очень хорошо рыболова, но не видит его. Это как раз из-за преломления луча зрения, описанного выше. Поэтому в зоне видимости маскировка имеет смысл. Следовательно, не стоит одевать на рыбалку одежду с яркими цветами, а наоборот в качестве маскировки подбирать более защитный цвет, который будет сливаться с общим фоном.
На мелководье вероятность того, что рыба заметит рыболова намного меньше, чем при ловле на более глубоких местах, вблизи берега. Из этого всего можно сделать вывод : что сидеть всегда лучше, чем стоять и меньше вероятности попасть в поле зрения рыбы. Вот почему и спиннингисту, который охотится с лодки, рекомендуется ловить рыбу (забрасывать приманку и вываживать хищника) сидя не только с целью соблюдения техники безопасности, но и стараться не быть замеченным рыбой.
Парные глаза, служащие органом зрения, обычно шаровидной формой и помещаются в глазных впадинах черепа, или орбитах. Снаружи каждый глаз защищен слоем хряща или плотной соединительной ткани, которые образуют толстую наружную оболочку глаза-склеру, или склеротик у (tunica fibrosa oculi; рис., II ). На передней открытой стороне склера заменена тонкой прозрачной оболочкой — роговицей (cornea) (рис., 10). На продольном разрезе глаза можно ясно видеть, что выпуклость роговицы иная, чем у склеры, вследствие чего роговица образует как бы полусферу, заметно выступающую вперед на профиле очертаний склеры. Изнутри склеротику, или склеру, выстилает сосудистая оболочка (tunica vasculosa), образованная соединительной тканью, обильно снабженной кровеносными сосудами.
К внешней стороне сосудистой оболочки иногда прилегает блестящая прослойка-зеркальце (tapetum lucidum), которое обусловливает собой свечение глаза в темноте. Обычно tapeturn lucidum слагается из нескольких клеточных слоев, между которыми залегают небольшие кристаллики, способные отражать световые лучи на лежащую кнутри от сооудистой оболочки сетчатку, или ретину. Такую структуру имеет tapetum lucidum у хищных млекопитающих (например, у кошек, собак), а также у китов и тюленей.
На месте соединения склеры и роговицы расположена кольцевая перепонка- радужина (iris; рис., 9). Соединительнотканный передний слой радужины может содержать пигмент, обусловливающий окраску темных глаз. Если пигмент в переднем слое совершенно отсутствует, но залегает в заднж слоях радужины, получается светлая окраска глаз (например, голубых).
Рис. Глаз щуки в вертикальном продольном разрезе.
1-сетчатка; 2-пигментный эпителий; 3 -сосудистая оболочка; 4- зрительный нерв; 5 — хрусталик; 4-его связка; 7-мышца, оттягиваюшяя хрусталик; 8-ее сухожилие; 9-радужина; 10 -роговица; 11-склеротика; 12-хориоидальная железа. Пунктиром показано положение хрусталика в аккомодированном глазу.
Наконец, если пигмента в радужине не содержится, просвечивающие кровеносные сосуды сообщают глазу красную расцветку, характерную для альбиносов. Радужина ограничивает внутреннее отверстие-зрачок, который может расширяться и сужаться благодаря наличию в слое радужины радиальных и кольцевых мускульных волокон. Непосредственно позади радужины образуется складчатый кольцевой валик-ресничное тело (corpus ciliare), состоящее из мускульных волокон. Это образование у наземных позвоночных имеет большое значение в процессе аккомодации или приспособления глаза к различному зрению (далекому и близкому).
Ресничное тело формируется из передних частей сосудистой оболочки. Кроме того, часть ретины, прилегающая к ресничному телу, образует зону мельчайших зубчиков (оrа serrata). Так, например, у человека имеется 50 подобных зубчиков. По краю ресничного тела прикрепляется с помощью соединительнотканных тяжей (zonula ciliaris zinnii) тонкая, также соединительнотканная сумка хрусталика. Прозрачный хрусталик (lens cristallina), состоящий из слоя тончайших концентрически расположенных волокон, в спокойном состоянии растянут связкой, посредством которой он прикреплен, и имеет относительно уплощенную форму (установка на далекое ); при сокращении ресничной мышцы ослабляется натяжение связки, и хрусталик становится более выпуклым (установка на близкое ).
У рыб аккомодация достигается наличием особого серповидного отростка, отходящего от сосудистой оболочки и прикрепляющегося к стенке хрусталика. Нежная полупрозрачная сетчатка, или ретина (retina; рис., I ), выстилает изнутри сосудистую оболочку. Края ретины доходят спереди до ресничного тела и заканчиваются здесь особыми волнистыми складками (см. выше). Зрительный нерв проходит с тыльной стороны глаза через склеру, сосудистую оболочку и своими волокнами распространяется по внутренней стороне сетчатки (рис., 4).
Строение ретины весьма сложно (рис. 2). Чувствительные клетки распределены на наружной (обращенной от света) стороне и несут каждая на своей наружной части особые воспринимающие элементы-относительно вытянутые палочки или более короткие вздутия-к о л б о ч к и (рис. 2, 9). Эти своеобразные образования погружаются концами в лежащий кнаружи пигментный слой ретины (рис. 2,10). Чувствительные клетки с описанными: воспринимающими элементами (палочками и колбочками) называются наружным клеточным слоем. Кнутри от него расположено несколько слоев нервных клеток, соединяющихся дендритами; к самому внутреннему слою ганглиозных клеток подходят окончания зрительного нерва (рис. 2, 12). Все эти сложные образования поддерживаются опорными клетками (рис. 2, 11).
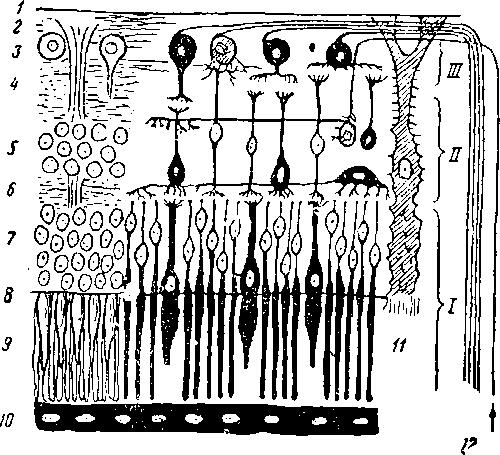
К описанию сетчатки следует добавить, что в месте вхождения в ретину зрительного нерва имеется лишь слой зрительных нервных волокон и отсутствуют элементы, воспринимающие свет. Участок сетчатки, соответствующий месту хождения зрительного нерва, не способен к восприятию световых раздражений и называется слепым пятном (papilla nervi optici). На линии главной оси глаза располагается желтое пятно (macula lutea; у птиц бывает до трех подобных пятен). На месте упомянутого пятна сетчатка состоит исключительно из слоя палочек и колбочек, т. е. наиболее восприимчива к свету.
Рис. 2. Схематический разрез через сетчатку глаза.
1-внутренняя пограничная перепонка; 2 -спой нервных волокон; 3 -слой ганглиозных клеток; 4 -внутренний сетчатый слой; 5 -слой биполярных клеток; 6 -наружный сетчатый слой; 7 -слой зрительных клеток; 8 -наружная пограничная перепонка; 9 -палочки и колбочки; 10 -пигментный эпителий; 11 -опорная ; 12 — центробежные и центростремительные волокна эрительного тракта; I, II, III -три последовательных пояса нейронов сетчатки.
Пространство между роговицей и зрачком называется передней камерой глаза и заполнено водянистой жидкостью. Небольшая узкая область между радужиной и хрусталиком составляет заднюю глазную камеру. Внутренняя полость глаза (между зрачком и ретиной) выполнена студенистым стекловидным телом (corpus vj treum), одетым тончайшей прозрачной бесструктурной оболочкой (membrana hyaloidea).
Роговица, водянистая жидкость, хрусталик и стекловидное тело составляют оптический аппарат глаза, служащий для проведения световых лучей к воспринимающему слою-сетчатке. При этом радужина служит диафрагмой, регулирующей количество света, поступающего в глаз. Одним из самых существенных отделов глаза является сетчатка, воспринимающая световые раздражения. У многих позвоночных глаз защищен складками кожи-в ерхним и нижним веком (palpebrae), выстланными изнутри многослойным эпителием, образующим конъюнктиву. В веках проходит кольцевая мускулатура, сокращение которой обусловливает сжимание и сближение век. По краям век часто можно видеть волосы в виде оторочек, образующих ресницы. У рыб имеется обычно только одно кольцевое веко. У некоторых млекопитающих, у птиц, у акул, амфибий и рептилий развита мигательная перепонка (membrana nicticans), лежащая глубже век и задергивающая глаза от внутреннего угла к наружному. У человека рудимент мигательной перепонки сохраняется в виде полулунной складки в нижнем внутреннем углу глаза. У многих ящериц и у рыб оба века прозрачны, сращены между собой и образуют постоянное прикрытие роговицы.
У наземных позвоночных в области век развиваются различные железы, смачивающие своими выделениями переднюю стенку глаза. Одна значительная группа желез локализована в области нижнего века у передне-внутреннего угла глаза,-здесь находится гардерова железа, выделяющая жирный секрет. Другая группа желез-так называемых слезных-находится у заднего угла глаза. У млекопитающих слезные железы расположены под верхним веком. Жидкое водянистое содержимое слезных желез стекает по конъюнктиве к передне-внутреннему углу глаза, откуда через слезный канал попадает в носовую полость. У водных млекопитающих, например, у китов, слезные железы хорошо развиты, но секрет их не жидкий, водянистый, а жирный и покрывает тонким слоем поверхность глазного яблока. Слезно-носовые каналы у китов не развиты.
Глаз позвоночных приводится в движение сокращениями шести мускулов. Из них четыре принадлежат к группе прямых мышц (m. recti). Внешний прямой мускул (см. выше) иннервируется n. abducens, три остальных-n. oculomotorius. Косых мышц две; из них верхняя снабжается окончаниями n. trochlearis, нижняя-n. oculomotorius. У многих млекопитающих, за исключением приматов, летучих мышей, слонов и др., развивается мускул, выпячивающий глаз (m. retractor bulbi). Этот мускул снабжается нервами: n. abducens, а в некоторых случаях и n. oculomotorius.