Для нормальной работы скелетных мышц необходимы ионы. Спортивное питание и здоровье Для работы скелетных мышц необходимы ионы
Минеральные вещества входят в состав всех живых тканей. Однако нормальное функционирование тканей обеспечивается не только наличием в них тех или иных минеральных солей, но и строго определенным их соотношением. Минеральные вещества поддерживают необходимое осмотическое давление в биологических жидкостях и обеспечивают постоянство кислотно-щелочного равновесия в организме.Рассмотрим основные минеральные вещества.
Калий содержится главным образом в клетках, натрий — в межклеточной жидкости. Для нормальной жизнедеятельности организма требуется строго определенное соотношение частиц натрия и калия. Должное соотношение этих ионов обеспечивает нормальную возбудимость нервной и мышечной тканей. Натрий играет большую роль в поддержании постоянства осмотического давления. При пониженном содержании калия в миокарде (мышечной ткани сердца) нарушается сократительная функция сердца. Но при избытке калия деятельность сердца также нарушается. Суточная потребность взрослого человека: натрий — 4-6 г, калий — 2-3 г.
Кальций входит в состав костей в виде фосфорных солей. Его ионы обеспечивают нормальную деятельность мозга и скелетных мышц. Наличие кальция необходимо для свертывания крови. Избыток кальция повышает частоту и силу сердечных сокращений, а при сверхбольших концентрациях в организме может вызвать остановку сердца. Суточная потребность взрослого человека в кальции — 0,7-0,8 г.
Фосфор входит в состав всех клеток и межтканевых жидкостей. Он играет большую роль в обмене белков, жиров, углеводов и витаминов. Это вещество — непременная составляющая богатых энергией веществ. Соли фосфорных кислот поддерживают постоянство кислотно-щелочного равновесия крови и других тканей. Суточная потребность взрослого человека в фосфоре — 1,5-2 г.
Хлор содержится в организме главным образом в соединении с натрием и входит в состав соляной кислоты желудочного сока. Хлор необходим для нормальной жизнедеятельности клеток. Суточная потребность взрослого человека в хлоре — 2-4 г.
Железо является составной частью гемоглобина и некоторых ферментов. Обеспечивая транспорт кислорода, оно принимает участие в окислительных процессах. Суточная потребность в железе для мужчин составляет 10 мг, для женщин — 18 мг.
Бром в небольших количествах содержится в крови и в других тканях. Усиливая торможение в коре больших полушарий, он способствует нормальному соотношению между процессами возбуждения и торможения.
Йод — обязательный компонент гормона щитовидной железы. Недостаток этого вещества в организме вызывает нарушение многих функций. Суточная потребность в йоде для взрослых здоровых людей составляет 0,15 мг (150 мкг).
Сера входит в состав многих белков. Она содержится в некоторых ферментах, гормонах, витаминах и других соединениях, играющих важную роль в обмене веществ. Кроме того, серная кислота используется печенью для нейтрализации некоторых веществ.
Для нормальной жизнедеятельности организма, кроме перечисленных веществ, имеют значение магний, цинк и т. д. Некоторые из них (алюминий, кобальт, марганец и др.) входят в состав организма в столь незначительных количествах, что их называют микроэлементами. Разнообразное питание обычно полностью обеспечивает организм всеми минералами.
Длинные тонкие мышечные волокна, из которых построена скелетная мышца, – это гигантские клетки, образующиеся в ходе онтогенеза при слиянии множества отдельных клеток. У взрослого человека они могут достигать в длину 5 см! Многочисленные ядра в такой клетке располагаются прямо под цитоплазматической мембраной, а основная часть цитоплазмы состоит из вытянутых вдоль всей клетки миофибрилл (толщиной 1–2 мкм), с характерной поперечной исчерченностью (рис. 3). Такую «раскраску» миофибрилле придают саркомеры, в состав каждого из которых входит два набора параллельных, частично перекрывающихся филаментов: толстых миозиновых, которые образуют темную полосу и тянутся от одного края до другого, и тонких актиновых, лежащих в области светлой полосы и частично заходящих в область темных полос (рис. 4). Один саркомер от другого отделяется Z-диском.
Рис. 3. Схема небольшого отрезка клетки скелетной мышцы (мышечного волокна)
Рис. 4. А. Электронная микрофотография продольного среза через клетку скелетной мышцы кролика (при малом увеличении). Видна регулярная поперечная исчерченность. Клетка содержит множество параллельных миофибрилл (см. рис. 3). Б. Небольшой участок того же фото: показаны отрезки двух смежных миофибрилл и детали саркомера. В. Схема строения отдельного саркомера, объясняющая происхождение темных и светлых полос, которые видны на электронной микрофотографии
Цитоплазма обеспечивает миофибриллы энергией в виде АТФ – в активно функционирующей мышце обнаруживается много митохондрий. Кроме того, в цитоплазме содержатся гликоген, фосфокреатин, гликолитические ферменты.
Скелетная мышца превращает химическую энергию, накопленную в АТФ, в механическую с очень большой эффективностью – в виде тепла теряется всего 30–50%. (Для сравнения: автомобильный двигатель при сжигании бензина обычно теряет в виде тепла 80–90%.)
При мышечном сокращении за счет связывания миозином актина толстые и тонкие нити скользят относительно друг друга (рис. 5).
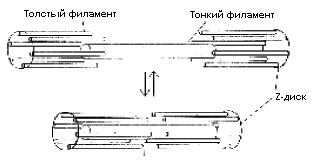
Рис. 5. Схема, иллюстрирующая процесс мышечного сокращения по принципу скользящих нитей: толстые и тонкие филаменты скользят друг по другу, не изменяя собственной длины
Физиологическим регулятором сокращения мышц служат ионы кальция. В состоянии покоя работает система активного транспорта ионов кальция, и они накапливаются в своеобразном хранилище, из которого высвобождаются под действием нервного импульса, обеспечивая мышечное сокращение.
Система транспорта ионов кальция работает за счет энергии АТФ. Того количества АТФ, которое имеется в мышце, хватает на поддержание работы сократительного аппарата всего в течение доли секунды. Как же работает мышца более продолжительное время? Оказывается, в мышце энергия запасается в форме фосфокреатина, или креатинфосфата, который может переносить больше высокоэнергетических фосфатных групп, чем универсальный АТФ. Фосфокреатин восстанавливает АТФ, обеспечивая тем самым приток энергии для мышечного сокращения. Однако в работающей мышце запасы фосфокреатина быстро истощаются, а это снижает и содержание АТФ.
При более продолжительной физической нагрузке мышцы обеспечиваются энергией за счет гликолиза – расщепления углеводов под действием ферментов с накоплением энергии в виде АТФ. Когда запасы креатина в мышце истощаются, понижается энергетический заряд мышечного сокращения, что приводит к стимуляции гликолиза, цикла трикарбоновых кислот и окислительного фосфорилирования в работающей мышце.
В отсутствие кислорода при расщеплении молекулы углеводов образуются две молекулы молочной кислоты (или лактата) и две молекулы АТФ. Однако, если для гликолиза используется гликоген мышц, то возникают две молекулы лактата и три молекулы АТФ.
Гликоген – это, как известно, главный резервный полисахарид, запасаемый в мышцах и печени. При пониженном уровне гликогена в мышцах и печени и наличии свободной глюкозы в крови она используется для синтеза гликогена. И, наоборот, при потребностях организма в энергетическом источнике для процессов гликолиза успешно используется гликоген.
Мышечная система наиболее развита по сравнению с другими системами организма. Для обеспечения работы мышц необходимо огромное количество энергии, которую человек может получать за счет креатинфосфата (фосфокреатина), углеводов в виде гликогена и глюкозы и жиров. Эти три вида энергоносителей различаются между собой по величине освобождаемой при их использовании энергии и по тому, как долго может каждый из них служить «топливным» источником. Так устроен механизм обеспечения мышц энергией (табл. 2, 3). Хорошо известно, что при продолжительной неинтенсивной работе при протекании окислительных процессов используются жиры или углеводы, а при работе несколько большей интенсивности используются механизмы анаэробного гликолиза. При очень интенсивной кратковременной нагрузке работа мышц обеспечивается за счет фосфагенов. Соответственно каждый из источников энергии имеет свою энергетическую стоимость и используется при определенных условиях (табл. 4).
Работающие мышцы для аэробного окисления углеводов по сравнению с другими органами потребляют очень большое количество кислорода (табл. 5).
Таблица 2. Запасы энергии в организме человека массой тела 70 кг
Таблица 3. Максимально возможная мощность скелетных мышц человека при использовании различных субстратов и путей катаболизма
Таблица 4. Максимальная скорость образования энергии из различных источников во время физических упражнений
|
Источник энергии |
Максимальная скорость мышцы |
Количество в мышце, ммоль/кг образования богатых энергией фосфатных связей, ммоль/ сек/кг |
Максимальная скорость продукции, ккал/ч/кг |
Время поддержания максимальной скорости |
|
Креатинфосфат |
||||
|
Анаэробный гликолиз |
||||
|
Аэробное окисление глюкозы и гликогена |
||||
|
Аэробное окисление |
Не лимитировано |
Не лимитировано |
Таблица 5. Относительное потребление кислорода различными органами человека в покое и при тяжелой работе*
Тонус скелетных мышц. В покое, вне работы, мышцы никогда полностью не расслабляются, а сохраняют некоторое напряжение, называемое тонусом. Внешне это выражается в упругости мышц. Тонус скелетных мышц связан с поступлением к мышце отдельных следующих друг за другом с большим интервалом нервных импульсов, возбуждающих попеременно различные мышечные волокна. Эти импульсы возникают в мотонейронах спинного мозга, активность которых в свою очередь поддерживается импульсами, исходящими как из вышележащих центров, так и с периферии от рецепторов растяжения («мышечных веретен»), находящихся в самих мышцах.
У человека тонус мышц в известных пределах может регулироваться произвольно: по желанию человек может почти полностью расслабить мышцы или же несколько напрячь их, не совершая, однако, при этом движения.
Работа и сила мышц. Величина сокращения (степень укорочения) мышцы при данной силе раздражения зависит как от ее морфологических свойств, так и от физиологического состояния. Длинные мышцы сокращаются на большую величину, чем короткие. Умеренное растяжение мышцы увеличивает ее сократительный эффект, при сильном же растяжении сокращение мышцы ослабляется. Если в результате длительной работы развивается утомление мышцы, то величина ее сокращения падает.
Для измерения силы мышцы определяют тот максимальный груз, который она в состоянии поднять. Эта сила может быть очень велика: собака, например, мышцами челюсти может поднять груз, превышающий вес ее тела в 8,3 раза. О силе икроножных мышц человека судят по величине груза, положенного ему на плечи, с которым он в состоянии приподняться на носки.
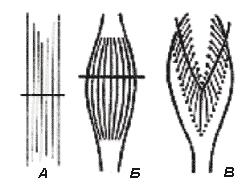
Рис. 6. Типы строения различных мышц (по А.А. Ухтомскому). А – мышцы с параллельным ходом волокон; Б – веретенообразная мышца; В – перистая мышца
Сила мышцы при прочих равных условиях зависит не от ее длины, а от поперечного сечения: чем больше физиологическое поперечное сечение мышцы, т.е. сумма поперечных сечений всех ее волокон, тем больше тот груз, который она в состоянии поднять. Физиологическое поперечное сечение совпадает с геометрическим только в мышцах с продольно расположенными волокнами; у мышц с косыми волокнами сумма поперечных сечений волокон может значительно превышать геометрическое поперечное сечение самой мышцы (рис. 6). По этой причине сила мышцы с косыми волокнами значительно больше, чем сила мышцы той же толщины, но с продольными волокнами. Чтобы сравнить силу разных мышц, максимальный груз, который мышца в состоянии поднять, делят на площадь ее поперечного сечения, вычисляя абсолютную мышечную силу. Например, абсолютная сила икроножной мышцы человека равна 5,9 кг/см 2 , сгибателя плеча – 8,1, жевательной мышцы – 10, двуглавой мышцы плеча – 11,4, трехглавой мышцы плеча – 16,8, гладких мышц – 1 кг/см 2 .
Большинство мышц млекопитающих и человека имеет перистое строение. Перистая мышца имеет большое физиологическое сечение, а поэтому обладает большой силой.
Работа мышцы измеряется произведением поднятого груза на величину укорочения мышцы, т.е. выражается в килограммометрах или граммсантиметрах.
По содержанию миоглобина мышечные волокна разделяют на красные, белые и промежуточные. Красные волокна принято считать «медленными», а белые – «быстрыми». Некоторые ученые считают, что при рождении мышцы человека состоят только из «медленных» волокон и часть из них в процессе развития превращается в «быстрые». Другие убеждены, что расположение мышц, особенности их строения и функций предопределены генетически.
Красные волокна работают в основном в аэробном режиме, а белые – в условиях недостатка кислорода, т.е. в них протекают разные метаболические процессы. Красные волокна, как правило, используются для выполнения легкой или умеренной работы, а белые начинают функционировать лишь после значительного возрастания притока возбуждающих импульсов во время очень интенсивной работы. Волокна промежуточного типа сохраняют свойства и красных, и белых волокон и называются еще «быстрыми красными».
Процентное содержание тех или иных волокон предопределяет специализацию атлета. Как правило, обладатели преимущественно красной мускулатуры достигают лучших результатов в видах спорта, требующих повышенной выносливости (плавание, велосипедный спорт, бег на средние и длинные дистанции и т.д.). Те, у кого больше белых мышечных волокон, лучше выполняют силовые упражнения. Последнее объясняется и тем, что белые волокна легче гипертрофируются (увеличиваются в объеме). Однако не все предопределяется только природой. Существуют еще и тренировочные факторы, которым некоторые специалисты отдают предпочтение в формировании структуры мышц.
Как мышцы «привыкают» к физическим нагрузкам. Во время тренировок в мышцах формируются механизмы запасания и использования энергетических субстратов: креатинфосфата, гликогена и жиров (в виде триацилглицеридов). Запасов АТФ и креатинфосфата, или фосфагена, в мышце очень мало. Фосфаты постоянно синтезируются (содержание креатинфосфата в белых мышечных волокнах составляет примерно 30 мкмоль на 1 г сырой массы мышцы), но быстро расходуются при нагрузке, продолжающейся более нескольких секунд. Энергоемкие гликоген и триацилглицериды составляют основную массу резервных источников «топлива» для мышц.
Под влиянием скоростной и силовой тренировки в белых мышечных волокнах повышается активность гликолитических ферментов и образуется большое количество гликогена, который запасается в мышцах в виде гранул, вокруг которых размещаются соответствующие ферменты. Происходят изменения и в буферных системах мышечных клеток: как только внутри клетки при интенсивной работе изменяется рН, это сразу же сказывается на работе ферментов, отвечающих за гликолиз.
Общее количество работы, которое анаэробный гликолиз может обеспечить при интенсивной нагрузке, зависит от резерва гликогена (при распаде каждой его молекулы образуется 6,2 моль АТФ). Само использование запасов гликогена в мышцах запускается гормональными и нервными стимулами. Один из таких гормонов – адреналин – способен значительно активизировать процессы использования гликогена для ресинтеза АТФ.
Под влиянием тренировки анаэробный гликолиз в мышцах можно многократно увеличить. Так, по некоторым данным, у тренированных спринтеров процессы гликолиза в мышцах ног усиливаются в две тысячи раз.
Однако за счет анаэробного гликолиза человек способен выполнять нагрузку только 2–3 минуты. После этого неизбежно запускаются процессы окислительного фосфорилирования.
При длительной работе главными действующими лицами становятся красные и промежуточные мышечные волокна. Энергия для деятельности этих мышц образуется в митохондриях (их в клетках красных мышечных волокон гораздо больше, чем в белых) с помощью окислительных ферментов в присутствии достаточного количества кислорода.
Гликоген, который активно используется при кратковременной работе, является (вместе с жирами) основным эндогенным субстратом и при продолжительной нагрузке. Оба эти вида «топлива», особенно жиры, содержатся в виде запасов в красных и промежуточных волокнах. Жиры несколько уступают гликогену в эффективности выхода энергии на единицу потребляемого кислорода: при их окислении образуется 5,6 моль АТФ.
При окислительном фосфорилировании мышца может получать энергетические субстраты и из центральных депо (гликоген из печени и жир из жировой ткани) и даже использовать энергетические источники, поступающие извне во время работы, например углеводные добавки при марафонском забеге, – природа предусмотрела для длительной работы дополнительные возможности.
Для представителей видов спорта, которые тренируются на выносливость, будет любопытен тот факт, что длительная и интенсивная работа мышц обеспечивается энергией лучше всего при одновременном использовании углеводов и жиров. Казалось бы, здесь некий парадокс. Ведь углеводов хватает только на 20–30 мин интенсивной работы, а жиры могут использоваться гораздо дольше. Однако дело все в том, что использование одних лишь жиров обеспечивает вдвое меньшую скорость выработки энергии, чем одновременное использование жиров и углеводов. А от этого зависит интенсивность выполняемой работы. Таким образом, как считают биохимики, гликоген является наилучшим «топливом» для обеспечения высокоинтенсивной продолжительной работы в аэробных условиях. Ученые нашли даже зависимость длительности работы до полного изнеможения от содержания гликогена в мышцах перед началом нагрузки. Однако, если нагрузка продолжается 2–3 ч, то организм начинает использовать для обеспечения мышечных сокращений и гликоген, и жиры. При переходе на расщепление жиров мощность работы снижается. Вначале используются триацилглицериды, а затем свободные жирные кислоты, которые поступают из крови.
Атлетов, специализирующихся в силовых видах спорта, и культуристов, конечно же, интересует вопрос рабочей гипертрофии мышц.
Систематическая интенсивная работа мышцы приводит к увеличению массы мышечной ткани. Это явление названо рабочей гипертрофией мышцы. В основе гипертрофии лежит увеличение массы протоплазмы мышечных волокон, приводящее к их утолщению. При этом повышается содержание белков и гликогена, а также веществ, доставляющих энергию, необходимую для мышечного сокращения – АТФ и креатинфосфата. Поэтому сила и скорость сокращения гипертрофированной мышцы выше, чем негипертрофированной.
Увеличение массы мышечной ткани у тренированных людей приводит к тому, что мускулатура тела может составлять 50% веса тела (вместо обычных 35–40%).
Гипертрофия развивается, если человек ежедневно на протяжении длительного времени производит мышечную работу, требующую большого напряжения (силовая нагрузка). Мышечная работа, производимая без особых усилий, даже если она продолжается очень долго, к гипертрофии мышцы не приводит.
Гипертрофия мышц очень важна для выполнения кратковременной «взрывной» работы. Возможно, это связано с тем, что запасы креатинфосфата в мышцах человека не увеличиваются больше определенного количества на единицу массы мышц. Таким образом, увеличение объема мышц способствует увеличению общего количества этого энергоемкого субстрата в мышцах и, соответственно, увеличению способности более эффективно выполнять работу максимальной мощности.
Противоположным рабочей гипертрофии явлением служит атрофия мышцы от бездеятельности. Она развивается во всех случаях, когда мышца почему-либо утрачивает способность совершать свою нормальную работу, например, при длительном обездвиживании конечности в гипсовой повязке, при долгом пребывании больного в постели, при перерезке сухожилия, вследствие чего мышца перестает совершать работу против нагрузки, и т.д.
При атрофии диаметр мышечных волокон и содержание в них сократительных белков, гликогена, АТФ и других важных для сократительной деятельности веществ резко падают. При возобновлении нормальной работы мышцы атрофия постепенно исчезает.
Утомление мышц. Временное понижение работоспособности мышц, наступающее в результате работы или тренировки и исчезающее после отдыха, определяется утомлением.
Если длительно раздражать ритмическими электрическими стимулами изолированную мышцу, к которой подвешен небольшой груз, то амплитуда ее сокращений постепенно убывает, пока не дойдет до нуля. Полученная таким образом кривая называется кривой утомления.
Наряду с изменением амплитуды сокращений при утомлении нарастает латентный период сокращения и понижается возбудимость. Но все эти изменения возникают не тотчас же после начала работы мыш-цы – существует некоторый период, в течение которого наблюдаются увеличение амплитуд сокращений и небольшое повышение возбудимости мышцы. При этом мышца становится легко растяжимой. В таких случаях говорят, что мышца «врабатывается», т.е. приспосабливается к работе при заданном ритме и силе раздражения. При дальнейшем длительном раздражении наступает утомление мышечных волокон.
Это может быть обусловлено накоплением в мышце продуктов обмена веществ (в частности, молочной кислоты, образующейся при расщеплении гликогена), оказывающих угнетающее влияние на работоспособность мышечных волокон. Часть этих продуктов, а также ионы калия диффундируют из волокон наружу в околоклеточное пространство и оказывают угнетающее влияние на способность возбудимой мембраны генерировать потенциалы действия.
Кроме того, на развитие утомления в мышце влияет постепенное истощение в ней энергетических запасов.
Все описанное выше относится к изолированной мышце. Ведь при работе в организме мышца непрерывно снабжается кровью и, следовательно, получает определенное количество питательных веществ (глюкозу, аминокислоты) и освобождается от продуктов обмена, нарушающих нормальную жизнедеятельность мышечных волокон. Главное отличие состоит в том, что в организме возбуждающие импульсы приходят к мышце с нерва. Нервно-мышечное соединение утомляется значительно раньше, чем мышечные волокна, в связи с чем блокирование передачи возбуждения с нерва на мышцу предохраняет последнюю от истощения, вызываемого длительной работой. В целостном организме еще раньше нервно-мышечных соединений утомляются при работе нервные центры.
Восстановление работоспособности утомленных мышц руки человека после длительной работы по подъему груза можно ускорить, если во время отдыха производить работу другой рукой или нижними конечностями.
Продолжение следует
На вопрос Чем обусловлено появление кальция в цитоплазме клеток скелетных мышц? заданный автором роскошествовать лучший ответ это кальций является фактором, разрешающим сокращение мышц: при повышении концентрации ионов кальция. в миоплазме происходит присоединение Сa к регуляторному белку, в результате чего актин становится способным взаимодействовать с миозином; соединяясь, эти два белка образуют актомиозин, и мышца сокращается. В процессе образования актомиозина происходит расщепление АТФ, химическая энергия которого обеспечивает выполнение механической работы и частично рассеивается в виде тепла. Наибольшая сократительная активность скелетной мышцы наблюдается при концентрации кальция 10-6-10(минус в) -7 моль; при понижении концентрации ионов Ca (менее 10-7 моль) мышечное волокно теряет способность к укорочению и напряжению. Действие Ca на ткани проявляется в изменении их трофики, интенсивности окислительно-восстановительных процессов и в других реакциях, связанных с образованием энергии. Изменение концентрации Ca в омывающей нервную клетку жидкости существенно влияет на проницаемость ее мембраны для ионов калия и особенно для ионов натрия, причем понижение уровня Ca вызывает повышение проницаемости мембраны для ионов натрия и повышение возбудимости нейрона. Повышение концентрации Ca оказывает стабилизирующее влияние на мембрану нервной клетки. Установлена роль Ca в процессах, связанных с синтезом и выделением нервными окончаниями медиаторов, обеспечивающих синаптическую передачу нервного импульса.
Перенос молекул и ионов против электрохимического градиента (активный транспорт) связан со значительными затратами энергии. Часто градиенты достигают больших величин. например, концентрационный градиент водородных ионов на плазматической мембране клеток слизистой оболочки желудка составляет 10в6степени, градиент концентрации ионов кальция на мембране саркоплазматического ретикулума — 10в4 степени, при этом потоки ионов против градиента значительны. В результате затраты энергии на транспортные процессы достигают, например, у человека, более 1/3 всей энергии метаболизма. В плазматических мембранах клеток различных органов обнаружены системы активного транспорта ионов натрия и калия — натриевый насос. Эта система перекачивает натрий из клетки и калий в клетку (антипорт) против их электрохимических градиентов. Перенос ионов осуществляется основным компонентом натриевого насоса — Na+, К+-зависимой АТФ-азой за счет гидролиза АТФ. На каждую гидролизующуюся молекулу АТФ транспортируется три иона натрия и два иона калия. Существуют два типа Са2+-АТФ-аз. Одна из них обеспечивает выброс ионов кальция из клетки в межклеточную среду, другая — аккумуляцию кальция из клеточного содержимого во внутриклеточное депо. Обе системы способны создавать значительный градиент иона кальция. К+, Н+-АТФ-аза обнаружена в слизистой оболочке желудка и кишечника. Она способна транспортировать Н+ через мембрану везикул слизистой оболочки при гидролизе АТФ. В микросомах слизистой оболочки желудка лягушки найдена аниончувствительная АТФ-аза, способная при гидролизе АТФ осуществлять антипорт бикарбоната и хлорида.
Сокращение мышц — это сложный процесс, состоящий из целого ряда этапов. Главными составляющими здесь являются миозин, актин, тропонин, тропомиозин и актомиозин, а также ионы кальция и соединения, которые обеспечивают мышцы энергией. Рассмотрим виды и механизмы мышечного сокращения. Изучим, из каких этапов они состоят и что необходимо для цикличного процесса.
Мышцы
Мышцы объединяются в группы, у которых одинаковый механизм мышечных сокращений. По этому же признаку они и разделяются на 3 вида:
- поперечно-полосатые мышцы тела;
- поперечно-полосатые мышцы предсердий и сердечных желудочков;
- гладкие мышцы органов, сосудов и кожи.
Поперечно-полосатые мышцы входят в опорно-двигательный аппарат, являясь его частью, так как помимо них сюда входят сухожилия, связки, кости. Когда реализуется механизм мышечных сокращений, выполняются следующие задачи и функции:
- тело передвигается;
- части тела перемещаются друг относительно друга;
- тело поддерживается в пространстве;
- вырабатывается тепло;
- кора активируется посредством афферентации с рецептивных мышечных полей.
Из гладких мышц состоит:
- двигательный аппарат внутренних органов, в который входят легкие и пищеварительная трубка;
- лимфатическая и кровеносная системы;
- система мочеполовых органов.
Физиологические свойства
Как и у всех позвоночных животных, в человеческом организме выделяют три самых важных свойства волокон скелетных мышц:
- сократимость — сокращение и изменение напряжения при возбуждении;
- проводимость — движение потенциала по всему волокну;
- возбудимость — реагирование на раздражитель посредством изменения мембранного потенциала и ионной проницаемости.
Мышцы возбуждаются и начинают сокращаться от идущих от центров. Но в искусственных условиях используют тогда может раздражаться напрямую (прямое раздражение) или через нерв, иннервирующий мышцу (непрямое раздражение).
Виды сокращений
Механизм мышечных сокращений подразумевает преобразование химической энергии в механическую работу. Этот процесс можно измерить при эксперименте с лягушкой: ее икроножную мышцу нагружают небольшим весом, а затем раздражают легкими электроимпульсами. Сокращение, при котором мышца становится короче, называется изотоническим. При изометрическом сокращении укорачивания не происходит. Сухожилия не позволяют при развитии укорачиваться. Еще один ауксотонический механизм мышечных сокращений предполагает условия интенсивных нагрузок, когда мышца укорачивается минимальным образом, а сила развивается максимальная.
Структура и иннервация скелетных мышц
В поперечно-полосатые скелетные мышцы входит множество волокон, находящихся в соединительной ткани и крепящихся к сухожилиям. В одних мышцах волокна расположены параллельно длинной оси, а в других они имеют косой вид, прикрепляясь к центральному тяжу сухожильному и к перистому типу.
Главная особенность волокна заключается в саркоплазме массы тонких нитей — миофибрилл. В них входят светлые и темные участки, чередующиеся друг с другом, а у соседних поперечно-полосатые волокна находятся на одном уровне — на поперечном сечении. Благодаря этому получается поперечная полосатость по всему волокну мышц.
Саркомером является комплекс из темного и двух светлых дисков, и он отграничивается Z-образными линиями. Саркомеры — это сократительный аппарат мышцы. Получается, что сократительное мышечное волокно состоит из:
- сократительного аппарата (системы миофибрилл);
- трофического аппарата с митохондриями, комплексом Гольджи и слабой ;
- мембранного аппарата;
- опорного аппарата;
- нервного аппарата.
Мышечное волокно разделяется на 5 частей со своими структурами и функциями и является целостной частью ткани мышц.
Иннервация
Этот процесс у поперечно-полосатых мышечных волокон реализуется посредством нервных волокон, а именно аксонов мотонейронов спинного мозга и головного ствола. Один мотонейрон иннервирует несколько волокон мышц. Комплекс с мотонейроном и иннервируемыми мышечными волокнами называют нейромоторной (НМЕ), или (ДЕ). Среднее число волокон, которые иннервирует один мотонейрон, характеризует величину ДЕ мышцы, а обратную величину называют плотностью иннервации. Последняя является большой в тех мышцах, где движения небольшие и «тонкие» (глаза, пальцы, язык). Малое ее значение будет, напротив, в мышцах с «грубыми» движениями (например, туловище).
Иннервация может быть одиночной и множественной. В первом случае она реализуется компактными моторными окончаниями. Обычно это характерно для крупных мотонейронов. (называющиеся в этом случае физическими, или быстрыми) генерируют ПД (потенциалы действий), которые распространяются на них.
Множественная иннервация встречается, к примеру, во внешних глазных мышцах. Здесь не генерируется потенциал действия, так как в мембране нет электровозбудимых натриевых каналов. В них распространяется деполяризация по всему волокну из синаптических окончаний. Это необходимо для того, чтобы привести в действие механизм мышечного сокращения. Процесс здесь происходит не так быстро, как в первом случае. Поэтому его называют медленным.
Структура миофибрилл
Исследования мышечного волокна сегодня проводятся на основе рентгеноструктурного анализа, электронной микроскопии, а также гистохимическими методами.
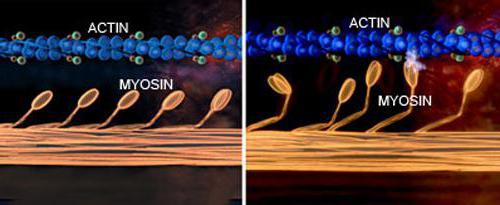
Рассчитано, что в каждую миофибриллу, диаметр которой составляет 1 мкм, входит примерно 2500 протофибрилл, то есть удлиненных полимеризованных молекул белков (актина и миозина). Актиновые протофибриллы в два раза тоньше миозиновых. В покое эти мышцы находятся так, что актиновые нити кончиками проникают в промежутки между миозиновыми протофибриллами.
Узкая светлая полоса в диске А свободна от актиновых нитей. А мембрана Z скрепляет их.
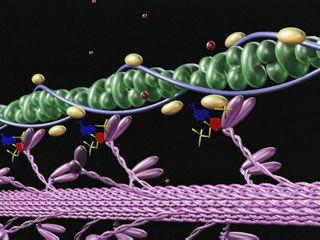
На миозиновых нитях есть поперечные выступы длиной до 20 нм, в головках которых находится порядка 150 молекул миозина. Они отходят биополярно, и каждая головка соединяет миозиновую с актиновой нитью. Когда происходит усилие актиновых центров на нитях миозина, актиновая нить приближается к центру саркомера. В конце миозиновые нити доходят до линии Z. Тогда они занимают собой весь саркомер, а актиновые находятся между ними. При этом длина диска I сокращается, а в конце он исчезает полностью, вместе с чем линия Z становится толще.
Так, по теории скользящих нитей, объясняется сокращение длины волокна мышцы. Теория, получившая название «зубчатого колеса», была разработана Хаксли и Хансоном в середине двадцатого века.
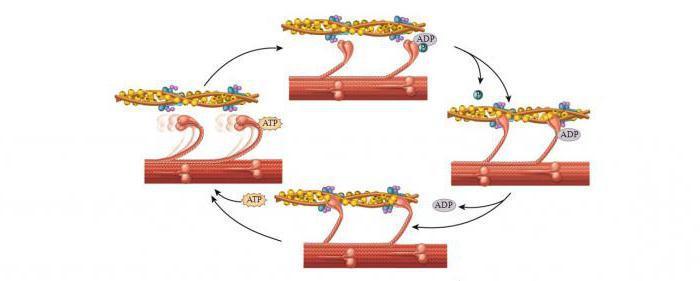
Механизм мышечного сокращения волокна
Главным в теории является то, что не нити (миозиновые и актиновые) укорачиваются. Длина их остается неизменной и при растяжении мышц. Но пучки тонких нитей, проскальзывая, выходят между толстыми нитями, уменьшается степень их перекрытия, таким образом происходит сокращение.
Молекулярный механизм мышечного сокращения посредством скольжения актиновых нитей заключается в следующем. Миозиновые головки соединяют протофибриллу с актиновой. При их наклонах происходит скольжение, двигающее актиновую нить к центру саркомера. За счет биполярной организации миозиновых молекул на обеих сторонах нитей создаются условия для скольжения актиновых нитей в разные стороны.
При расслаблении мышц миозиновая головка отходит от актиновых нитей. Благодаря легкому скольжению расслабленные мышцы растяжению сопротивляются гораздо меньше. Поэтому они пассивно удлиняются.
Этапы сокращения
Механизм мышечного сокращения кратко можно подразделить на следующие этапы:
- Мышечное волокно стимулируется, когда потенциал действия поступает от мотонейронов из синапсов.
- Потенциал действия создается на мембране мышечного волокна, а затем распространяется к миофибриллам.
- Совершается электромеханическое сопряжение, представляющее собой преобразование электрического ПД в механическое скольжение. В этом обязательно участвуют ионы кальция.
Ионы кальция
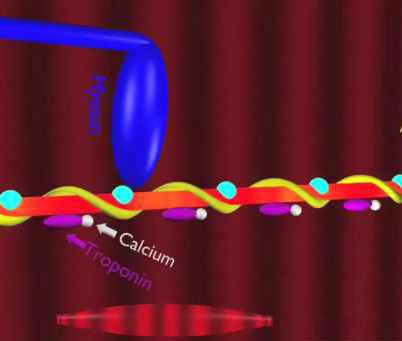
Для лучшего понимания процесса активации волокна ионами кальция удобно рассмотреть структуру актиновой нити. Длина ее составляет порядка 1 мкм, толщина — от 5 до 7 нм. Это пара закрученных ниток, которые напоминают мономер актина. Примерно через каждые 40 нм здесь находятся сферические тропониновые молекулы, а между цепями — тропомиозиновые.
Когда ионы кальция отсутствуют, то есть миофибриллы расслабляются, длинные тропомиозиновые молекулы блокируют крепление актиновых цепей и мостиков миозина. Но при активизации ионов кальция тропомиозиновые молекулы опускаются глубже, и участки открываются.
Тогда миозиновые мостики прикрепляются к актиновым нитям, а АТФ расщепляется, и сила мышц развивается. Это становится возможным за счет воздействия кальция на тропонин. При этом молекула последнего деформируется, проталкивая тем самым тропомиозин.
Когда мышца расслаблена, в ней на 1 грамм сырого веса содержится больше 1 мкмоль кальция. Соли кальция изолированы и находятся в особых хранилищах. В противном случае мышцы бы все время сокращались.
Хранение кальция происходит следующим образом. На разных участках мембраны клетки мышцы внутри волокна имеются трубки, через которые происходит соединение со средой вне клеток. Это система поперечных трубочек. А перпендикулярно ей находится система продольных, на концах которых — пузырьки (терминальные цистерны), расположенные в непосредственной близости к мембранам поперечной системы. Вместе получается триада. Именно в пузырьках хранится кальций.
Так ПД распространяется внутрь клетки, и происходит электромеханическое сопряжение. Возбуждение проникает в волокно, переходит в продольную систему, высвобождает кальций. Таким образом осуществляется механизм сокращения мышечного волокна.
3 процесса с АТФ
При взаимодействии обеих нитей при наличии ионов кальция немалая роль отводится АТФ. Когда реализуется механизм мышечного сокращения скелетной мышцы, энергия АТФ применяется для:
- работы насоса натрия и калия, который поддерживает постоянную концентрацию ионов;
- этих веществ по разные стороны мембраны;
- скольжения нитей, укорачивающих миофибриллы;
- работы насоса кальция, действующего для расслабления.
АТФ находится в клеточной мембране, нитях миозина и мембранах ретикулума саркоплазматического. Фермент расщепляется и утилизируется миозином.
Потребление АТФ
Известно, что миозиновые головки взаимодействуют с актином и содержат элементы для расщепления АТФ. Последняя активизируется актином и миозином при наличии ионов магния. Поэтому расщепление фермента происходит при прикреплении миозиновой головки к актину. При этом чем больше поперечных мостиков, тем скорость расщепления будет выше.
Механизм АТФ
После завершения движения молекула АФТ обеспечивает энергией для разделения участвующих в реакции миозина и актина. Миозиновые головки разделяются, АТФ расщепляется до фосфата и АДФ. В конце подсоединяется новая АТФ-молекула, и цикл возобновляется. Таковым является механизм мышечного сокращения и расслабления на молекулярном уровне.
Активность поперечных мостиков будет продолжаться лишь до тех пор, пока происходит гидролиз АТФ. При блокировке фермента мостики не станут снова прикрепляться.
С наступлением смерти организма уровень АТФ в клетках падает, и мостики остаются устойчиво прикрепленными к актиновой нити. Так происходит стадия трупного окоченения.
Ресинтез АТФ
Ресинтез возможно реализовать двумя путями.
Посредством ферментативного переноса от креатинфосфата фосфатной группы на АДФ. Так как запасов в клетке креатинфосфата намного больше АТФ, ресинтез реализуется очень быстро. В то же время посредством окисления пировиноградной и молочной кислот ресинтез будет осуществляться медленно.
АТФ и КФ могут исчезнуть полностью, если ресинтез будет нарушен ядами. Тогда и кальциевый насос прекратит работу, вследствие чего мышца необратимо сократится (то есть настанет контрактура). Таким образом, нарушится механизм мышечного сокращения.
Физиология процесса
Подытоживая вышесказанное, отметим, что сокращение волокна мышцы состоит в укорочении миофибрилл в каждом из саркомеров. Нити миозина (толстые) и актина (тонкие) связаны концами в расслабленном состоянии. Но они начинают скользящие движения друг навстречу к другу, когда реализуется механизм мышечного сокращения. Физиология (кратко) объясняет процесс, когда под влиянием миозина выделяется необходимая энергия для преобразования АТФ в АДФ. При этом активность миозина будет реализована лишь при достаточном содержании ионов кальция, накапливающихся в саркоплазматической сети.
Внутренних органов, кожи, сосудов.
Скелетные мышцы совместно со скелетом составляют опорно-двигательную систему организма, которая обеспечивает поддержание позы и перемещение тела в пространстве. Кроме того, они выполняют защитную функцию, предохраняя внутренние органы от повреждений.
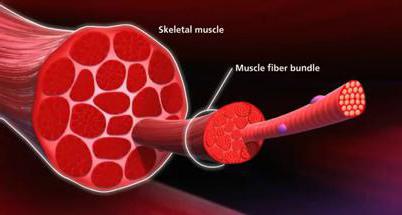
Скелетные мышцы являются активной частью опорно-двигательного аппарата, включающего также кости и их сочленения, связки, сухожилия. Масса мышц может достигать 50% общей массы тела.
С функциональной точки зрения к двигательному аппарату можно отнести и моторные нейроны, посылающие нервные импульсы к мышечным волокнам. Тела моторных нейронов, иннервирующих аксонами скелетную мускулатуру, располагаются в передних рогах спинного мозга, а иннервирующих мышцы челюстно-лицевой области — в моторных ядрах ствола мозга. Аксон мотонейрона при входе в скелетную мышцу ветвится, и каждая веточка участвует в формировании нервно-мышечного синапса на отдельном мышечном волокне (рис. 1).
Рис. 1. Разветвления аксона моторного нейрона на аксонные терминалы. Электронограмма
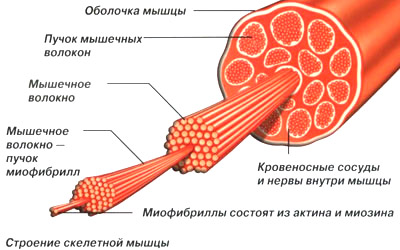
Рис. Строение скелетной мышцы человека
Скелетные мышцы состоят из мышечных волокон, которые объединяются в мышечные пучки. Совокупность мышечных волокон, иннервируемых веточками аксона одного моторного нейрона, называют двигательной (или моторной) единицей. В глазных мышцах 1 двигательная единица может содержать 3-5 мышечных волокон, в мышцах туловища — сотни волокон, в камбаловидной мышце — 1500-2500 волокон. Мышечные волокна 1 двигательной единицы имеют одинаковые морфофункциональные свойства.
Функциями скелетных мышц являются:
- передвижение тела в пространстве;
- перемещение частей тела относительно друг друга, в том числе осуществление дыхательных движений, обеспечивающих вентиляцию легких;
- поддержание положения и позы тела.
Скелетные мышцы вместе со скелетом составляют опорно-двигательную систему организма, которая обеспечивает поддержание позы и перемещение тела в пространстве. Наряду с этим скелетные мышцы и скелет выполняют защитную функцию, предохраняя внутренние органы от повреждения.
Кроме того, поперечно-полосатые мышцы имеют значение в выработке тепла, поддерживающего температурный гомеостаз, и в депонировании некоторых питательных веществ.
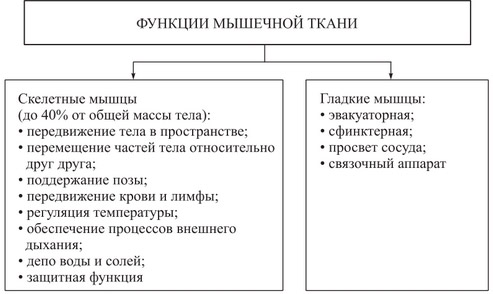
Рис. 2. Функции скелетных мышц
Физиологические свойства скелетных мышц
Скелетные мышцы обладают следующими физиологическими свойствами.
Возбудимость. Обеспечивается свойством плазматической мембраны (сарколеммы) отвечать возбуждением на поступление нервного импульса. Из-за большей разности потенциала покоя мембраны поперечно-полосатых мышечных волокон (Е 0 около 90 мВ) возбудимость их ниже, чем нервных волокон (Е 0 около 70 мВ). Амплитуда потенциала действия у них больше (около 120 мВ), чем у других возбудимых клеток.
Это позволяет на практике достаточно легко регистрировать биоэлектрическую активность скелетных мыши. Длительность потенциала действия составляет 3-5 мс, что определяет короткую продолжительность фазы абсолютной рефрактерности возбужденной мембраны мышечных волокон.
Проводимость. Обеспечивается свойством плазматической мембраны формировать локальные круговые токи, генерировать и проводить потенциал действия. В результате потенциал действия распространяются по мембране вдоль мышечного волокна и вглубь по поперечным трубочкам, формируемым мембраной. Скорость проведения потенциала действия составляет 3-5 м/с.
Сократимость. Представляет собой специфическое свойство мышечных волокон изменять свою длину и напряжение вслед за возбуждением мембраны. Сократимость обеспечивается специализированными сократительными белками мышечного волокна.
Скелетные мышцы обладают также вязкоэластическими свойствами, имеющими важное значение для расслабления мышц.
Рис. Скелетные мышцы человека
Физические свойства скелетных мышц
Скелетные мышцы характеризуются растяжимостью, эластичностью, силой и способностью совершать работу.
Растяжимость — способность мышцы изменять длину под действием растягивающей силы.
Эластичность — способность мышцы восстанавливать первоначальную форму после прекращения действия растягивающей или деформирующей силы.
— способность мышцы поднимать груз. Для сравнения силы различных мышц определяют их удельную силу путем деления максимальной массы на число квадратных сантиметров ее физиологического сечения. Сила скелетной мышцы зависит от многих факторов. Например, от числа двигательных единиц, возбуждаемых в данный момент времени. Также она зависит от синхронности работы двигательных единиц. Сила мышцы зависит и от исходной длины. Существует определенная средняя длина, при которой мышца развивает максимальное сокращение.
Сила гладких мышц тоже зависит от исходной длины, синхронности возбуждения мышечного комплекса, а также от концентрации ионов кальция внутри клетки.
Способность мышцы совершать работу. Работа мышцы определяется произведением массы поднятого груза на высоту подъема.
Работа мышц возрастаете увеличением массы поднимаемого груза, но до определенного предела, после которого увеличение груза приводит к уменьшению работы, т.е. снижается высота подъема. Максимальная работа совершается мышцей при средних нагрузках. Это называется законом средних нагрузок. Величина мышечной работы зависит от числа мышечных волокон. Чем толще мышца, тем больший груз она может поднять. Длительное напряжение мышцы приводит к ее утомлению. Это обусловлено истощением энергетических запасов в мышце (АТФ, гликоген, глюкоза), накоплением молочной кислоты и других метаболитов.
Вспомогательные свойства скелетной мускулатуры
Растяжимость — это способность мышцы изменять свою длину под действием растягивающей ее силы. Эластичность — способность мышцы принимать свою первоначальную длину после прекращения действия растягивающей или деформирующей силы. Живая мышца обладает малой, но совершенной эластичностью: уже небольшая сила способна вызвать относительно большое удлинение мышцы, а возвращение ее к первоначальным размерам является полным. Это свойство очень важно для осуществления нормальных функций скелетных мышц.
Сила мышцы определяется максимальным грузом, который мышца в состоянии поднять. Для сравнения силы различных мышц определяют их удельную силу, т.е. максимальный груз, который мышца в состоянии поднять, делят на число квадратных сантиметров ее физиологического поперечного сечения.
Способность мышцы совершать работу. Работа мышцы определяется произведением величины поднятого груза на высоту подъема. Работа мышцы постепенно увеличивается с увеличением груза, но до определенного предела, после которого увеличение груза приводит к уменьшению работы, так как снижается высота подъема груза. Следовательно, максимальная работа мышцей производится при средних величинах нагрузок.
Утомление мышц. Мышцы не могут работать беспрерывно. Длительная работа приводит к снижению их работоспособности. Временное понижение работоспособности мышцы, наступающее при длительной работе и исчезающее после отдыха, называется утомлением мышцы. Принято различать два вида утомления мышц: ложное и истинное. При ложном утомлении утомляется не мышца, а особый механизм передачи импульсов с нерва на мышцу, называемый синапсом. В синапсе истощаются резервы медиаторов. При истинном утомлении в мышце происходят следующие процессы: накопление недоокисленных продуктов распада питательных веществ вследствие недостаточного поступления кислорода, истощение запасов источников энергии, необходимой для мышечного сокращения. Утомление проявляется уменьшением силы сокращения мышцы и степени расслабления мышцы. Если мышца на некоторое время прекращает работу и находится в состоянии покоя, то восстанавливается работа синапса, а с кровью удаляются продукты обмена и доставляются питательные вещества. Таким образом, мышца вновь приобретает способность сокращаться и производить работу.
Одиночное сокращение
Раздражение мышцы или иннервирующего ее двигательного нерва одиночным стимулом вызывает одиночное сокращение мышцы. Различают три основные фазы такого сокращения: латентная фаза, фаза укорочения и фаза расслабления.
Амплитуда одиночного сокращения изолированного мышечного волокна от силы раздражения не зависит, т.е. подчиняется закону «все или ничего». Однако сокращение целой мышцы, состоящей из множества волокон, при ее прямом раздражении зависит от силы раздражения. При пороговой силе тока в реакцию вовлекается лишь небольшое число волокон, поэтому сокращение мышцы едва заметно. С увеличением силы раздражения число волокон, охваченных возбуждением, возрастает; сокращение усиливается до тех пор, пока все волокна не оказываются сокращенными («максимальное сокращение») — этот эффект называется лестницей Боудича. Дальнейшее усиление раздражающего тока на сокращение мышцы не влияет.
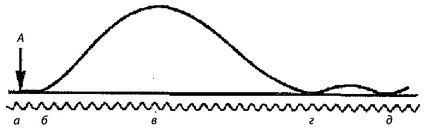
Рис. 3. Одиночное сокращение мышцы: А — момент раздражения мышцы; а-6 — скрытый период; 6-в — сокращение (укорочение); в-г — расслабление; г-д — последовательные эластические колебания.
Тетанус мышцы
В естественных условиях к скелетной мышце из центральной нервной системы поступают не одиночные импульсы возбуждения, которые служат для нее адекватными раздражителями, а серии импульсов, на которые мышца отвечает длительным сокращением. Длительное сокращение мышцы, возникающее в ответ на ритмическое раздражение, получило название тетанического сокращения, или тетануса. Различают два вида тетануса: зубчатый и гладкий (рис. 4).
Гладкий тетанус возникает, когда каждый последующий импульс возбуждения поступает в фазу укорочения, а зубчатый — в фазу расслабления.
Амплитуда тетанического сокращения превышает амплитуду одиночного сокращения. Академик Н.Е. Введенский обосновал изменчивость амплитуды тетануса неодинаковой величиной возбудимости мышцы и ввел в физиологию понятия оптимума и пессимума частоты раздражения.
Оптимальной называется такая частота раздражения, при которой каждое последующее раздражение поступает в фазу повышенной возбудимости мышцы. При этом развивается тетанус максимальной величины (оптимальный).
Пессимальной называется такая частота раздражения, при которой каждое последующее раздражение осуществляется в фазу пониженной возбудимости мышцы. Величина тетануса при этом будет минимальной (пессимальной).
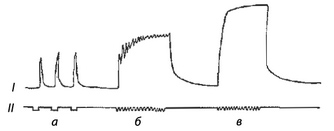
Рис. 4. Сокращение скелетной мышцы при разной частоте раздражения: I — сокращение мышцы; II — отметка частоты раздражения; а — одиночные сокращения; б- зубчатый тетанус; в — гладкий тетанус
Режимы мышечных сокращений
Для скелетных мышц характерны изотонический, изометрический и смешанный режимы сокращения.
При изотоническом сокращении мышцы изменяется ее длина, а напряжение остается постоянным. Такое сокращение происходит в том случае, когда мышца не преодолевает сопротивления (например, не перемещает груз). В естественных условиях близкими к изотоническому типу сокращениями являются сокращения мышц языка.
При изометрическом сокращении в мышце во время ее активности нарастает напряжение, но из-за того, что оба конца мышцы фиксированы (например, мышца пытается поднять большой груз), она не укорачивается. Длина мышечных волокон остается постоянной, меняется лишь степень их напряжения.
Сокращаются по аналогичным механизмам.
В организме сокращения мышц никогда не бывают чисто изотоническими или изометрическими. Они всегда имеют смешанный характер, т.е. происходит одновременное изменение и длины, и напряжения мышцы. Такой режим сокращения называется ауксотоническим, если преобладает напряжение мышцы, или ауксометрическим, если преобладает укорочение.